

 , 葛晓阳, 李付广, 张朝军
, Ge Xiaoyang, Li Fuguang, Zhang Chaojun
, 葛晓阳, 李付广, 张朝军
, Ge Xiaoyang, Li Fuguang, Zhang Chaojun
茉莉酸在调控棉花体细胞胚增殖方面有重要作用,而丙二烯氧化物合酶(AOS)是植物茉莉酸合成途径中的关键酶。作者鉴定出在胚发育过程中表达的AOS基因,这些基因编码区长度均在1500 bp左右,被定位在5条染色体上,有2个基因含有内含子。对其启动子元件分析发现,它们可能受激素和胁迫的调控在体细胞胚胎发生中起作用。荧光定量分析发现,这些基因在棉花不同组织中的表达模式主要分为3类,GhAOS1和GhAOS6在棉花胚发育过程中表达量相对较高,GhAOS2和GhAOS3在棉花愈伤和胚性愈伤中表达量相对较高,GhAOS4和GhAOS5在茎和叶中表达水平相对较高,暗示这些基因的表达有时空特异性,且GhAOS1和GhAOS6可能在棉花胚发育中发挥重要作用。亚细胞定位分析显示,大部分被定位在过氧化物酶体中,可能参与调控光呼吸、体细胞胚成熟和萌发。上述分析为进一步研究AOS基因家族在棉花体细胞胚发育中的功能奠定了基础。
Allene oxide synthase (AOS) is the key enzyme in the jasmonic acid (JA) biosynthesis pathway. As a class of plant hormones, JA plays an important role in the regulation of plant growth and development, but studies on JAs regulatory role in somatic embryo development are lacking. Here, we identified AOS family members during cotton somatic embryo development. The opening read frames of the six genes are all ~1500 bp, but only two genes contain introns, and the genes are dispersed across five different chromosomes. Based on the expression profile clusters, these genes are mainly divided into three subgroups: the high expression levels of GhAOS1 and GhAOS6 in cotton somatic embryos, the high expression levels of GhAOS2 and GhAOS3 in calli and embryogenetic calli, and the high expression levels of GhAOS4 and GhAOS5 in cotton stems and leaves.The groupings suggest that the genes play different roles during somatic embryo development. Most AOS genes were located in the peroxisome, which may be involved in photorespiration, and somatic embryo germination and maturation. The identification and bioinformatics analyses laid the foundation for the function analysis of the AOS gene family in cotton somatic embryos.
茉莉酸类化合物(Jasmonates,JAs)包括茉莉酸(Jasmonic acid,JA)、茉莉酸甲酯(Methyl Jasmonates,Me-JA)以及其他的衍生物等,是一类基本的植物激素[1]。JA作为一种新型植物激素,一方面调节植物的生长和发育等[1-2],另一方面作为内源信号分子参与生物胁迫防御和非生物胁迫应答的信号传递[3-6]。JA在植物体细胞胚胎发生方面也发挥着重要的作用。在大豆顶端分生组织中,JA浓度高于0.1 μmol·L-1时抑制愈伤组织的形成[7];在番茄顶端分生组织中,JA浓度在0.5~25 μmol·L-1时严重抑制了愈伤组织的形成[8];Me-JA能够推迟胡萝卜从胚性细胞到鱼雷胚的分化时间并抑制体细胞胚的再生[9]。
丙二烯氧化物合酶(Allene oxide synthase,AOS)是植物JA合成途径中的关键酶[10],与JA的生物合成关系密切。Hanns等[11]发现在马铃薯中转入亚麻AOS基因的cDNA,其内源JA含量比未转基因植株高8~12倍。AOS基因的研究在水稻、拟南芥、银胶菊和亚麻等作物中也都有报道:拟南芥叶片受到伤害后,其AOS基因转录水平上升,内源JA的含量升高来应对伤害产生的胁迫[12];水稻AOS家族共有4个成员,分别是OsAOS1、OsAOS2、OsAOS3和OsAOS4,这4个基因在不同组织中的表达模式不相同,在应对重金属和伤害诱导方面的响应也不相同,暗示这些基因发挥不同的作用[13-15];AOS在银胶菊和亚麻中的作用与在其他植物中相似,作为一个调控因子在抗逆方面起重要作用[16-17]。目前报道的关于AOS基因的研究都与伤害和逆境胁迫有关,而逆境胁迫如渗透压、重金属、温度、伤害、化学试剂等非生物胁迫以及生物胁迫都可以促进体细胞胚胎发生[18]。JA在调控体细胞胚胎发生中发挥着重要的作用,而已报道的AOS基因在伤害等逆境胁迫下表达水平发生变化,推测逆境胁迫调控AOS基因诱导内源JA含量发生变化,进而在体细胞胚胎发生中发挥作用。
利用本实验室的棉花体细胞胚发育转录组数据,挑选出AOS-like基因,与陆地棉基因组数据库进行序列比对,获得相应的DNA、CDS(Coding sequence)和推测的氨基酸序列。根据氨基酸序列,在NCBI上进行blastp比对,得到相似性较高的19个不同物种的AOS序列。利用MEGA 5.05软件对鉴定的棉花AOS蛋白和19个其他物种的AOS蛋白进行多种序列比对,采用邻接(Neighbor-Joining, NJ)算法进行系统进化树分析。
根据核苷酸序列在陆地棉染色体序列中进行比对,设定E≤10-5,选取序列相似度不小于98%的序列,得到其基因在染色体中的位置,用Mapchart 2.3软件绘制染色体定位图。利用PSORT软件(http://psort.hg c.jp/)对氨基酸序列分析,并进行亚细胞定位预测分析。
将AOS基因起始密码子上游的2000 bp序列作为潜在的启动子序列,在启动子作用 元 件 分 析 网 站 PLACE(http://www.dna.affrc.go.jp/ PLACE/)和PLANTCARE (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)上进行分析,预测可能存在的 TATA-box,CAAT-box以及其他可能被激素和压力诱导响应的顺式作用元件。
1.4.1 RNA提取与反转录。陆地棉中棉所24根、茎、叶、花、愈伤组织、胚性愈伤组织、球形胚、鱼雷胚、子叶胚总RNA的提取按照天根RNA提取试剂盒说明书,通过10 g·L-1的琼脂糖凝胶(0.2 g的琼脂糖溶解于20 mL的TAE电泳液)电泳和Nanodrop检测RNA纯度与浓度,取质量较好的RNA样本进行反转录,20 μL反应体系:5×Prime Script R Buffer (for Real-time) 4 μL,Prime Script R RT Enzyme MixⅠ 1 μL,Oligo dT Primer (50 μmol·L-1) 1 μL,Random 6 mers (100 μmol·L-1) 1 μL, RNase Free dH2O补至20 μL,总RNA为1 μg,具体步骤见TaRaKa One Step SYBR® PrimeScript ® RT-PCR Kit 说明书。
1.4.2 荧光定量qRT-PCR(Quantitative real-time PCR)。选用ABI7900仪器(美国)进行荧光定量qRT-PCR,分析鉴定出的陆地棉AOS基因家族成员在根、茎、叶、花、愈伤组织、胚性愈伤组织、球形胚、鱼雷胚、子叶胚中的表达情况,以GhHistone 3为内参基因,所用引物见
表1 实时荧光定量PCR引物序列
Table 1 Primer sequence used for quantitative RT-PCR
利用棉花体细胞胚发育转录组数据,将获得的unigenes利用GO、COG和KEGG数据库进行功能注释,获得6个候选的AOS同源基因。根据氨基酸和保守结构域的相似性,发现这6个基因所编码的氨基酸序列在C端都含有保守的p450保守结构域,与已经报道的AOS氨基酸序列有很高的相似性,确定候选基因属于AOS基因家族,分别命名为GhAOS1、GhAOS2、GhAOS3、GhAOS4、GhAOS5和GhAOS6。
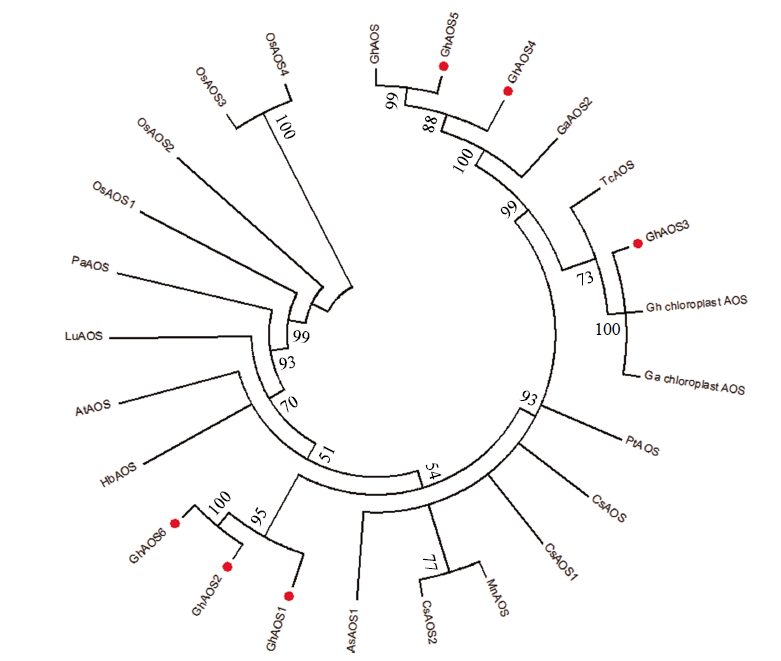
将预测出的6个AOS基因所编码的氨基酸序列与在NCBI中比对到不同物种的19个AOS氨基酸序列进行多重序列比对,构建系统进化树。根据进化树分析发现GhAOS1、GhAOS2和GhAOS6聚为一类,相似性较高,其中GhAOS2与GhAOS6的相似性为100%;而GhAOS3与陆地棉及亚洲棉叶绿体中存在的AOS相似度为100%;另外,GhAOS4与GhAOS5的相似性为88%,这两者与NCBI上能够查到的GhAOS(GenBank: ALG62633.1)序列聚为一类,GhAOS5与GhAOS相似性为99%(
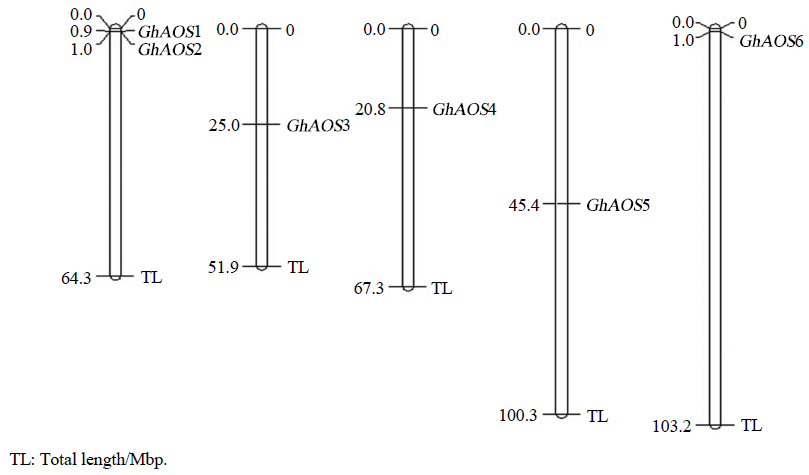
利用陆地棉AADD基因组数据库,对陆地棉AOS基因家族进行内含子和外显子分析,发现GhAOS2、GhAOS3、GhAOS4、GhAOS5都不存在内含子,CDS长度均在1500 bp左右;而GhAOS1和GhAOS6各有1个内含子,GhAOS1内含子长度为1455 bp,GhAOS6内含子长度为3223 bp。因此,从基因结构上,可将6个AOS分为2类,一类含有内含子,一类不含有内含子。染色体定位发现,6个基因被定位在5条染色体上,其中GhAOS1和GhAOS2被定位在同一条染色体上(
亚细胞定位预测结果表明:GhAOS1、GhAOS4、GhAOS5和GhAOS6都被定位在过氧化物酶体上,而GhAOS2被定位在线粒体基质内,GhAOS3被定位在叶绿体基质中(
表2 陆地棉AOS蛋白的亚细胞定位
Table 2 Subcellular localization of AOS proteins
利用棉花AADD基因组数据,对这6个AOS基因的起始密码子上游的2000 bp序列分析,发现它们均具有典型核心启动子TATA盒和CAAT盒,可以确认这些序列具有调控转录功能。除GhAOS3外,另5个基因具有响应Me-JA的调控元件CGTCA基序和TGACG基序,所有的AOS启动子都含有SKn-1基序和响应激素的顺式元件,GhAOS1的启动子存在1个响应脱落酸顺式元件ABRE;GhAOS2的启动子存在与乙烯,水杨酸和生长素调节相关的元件ERE、TCA和TGA;GhAOS3的启动子存在脱落酸和赤霉素响应元件ABRE和P-box;GhAOS4和GhAOS5的启动子都存在乙烯响应元件ERE;GhAOS6的启动子存在乙烯和水杨酸响应元件ERE和TCA。除了上述调控元件外,不同基因的启动子至少含有1个胁迫响应顺式元件(ARE、HSE、TC-rich Repeats、LTR或MBS),且GhAOS2和GhAOS6都存在与伤害诱导相关的WUN基序(
表3 陆地棉AOS基因家族启动子顺式作用元件预测
Table 3 The cis-acting elements of promoter on the G. hirsutum
依据转录组数据检测结果(
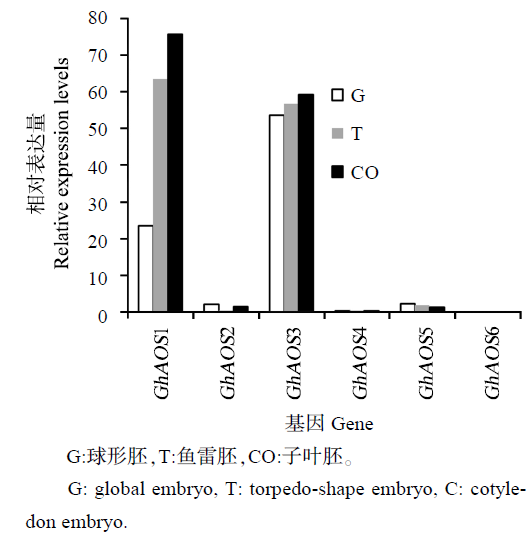
图4 陆地棉AOS基因家族在体细胞胚转录组中的相对表达量
Fig. 4 Relative expression levels of AOS genes based on somatic embryo transcription
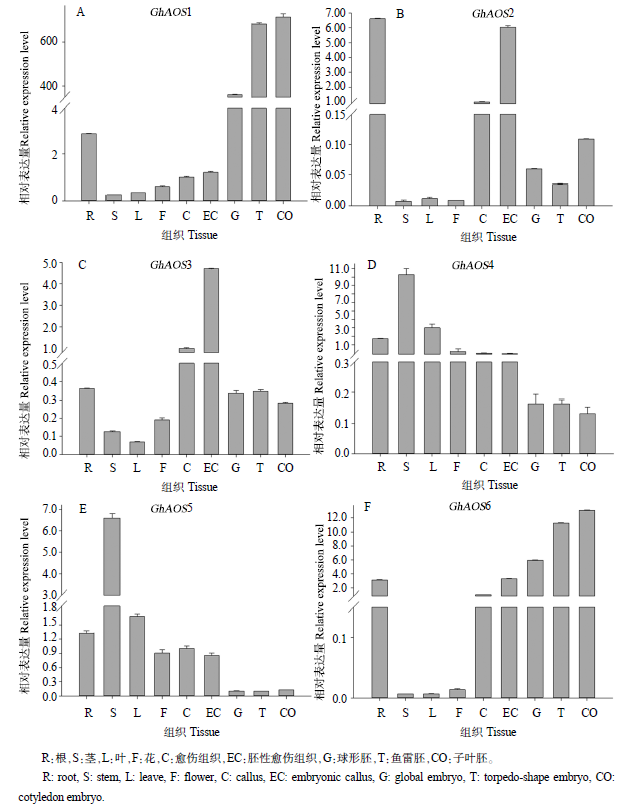
图5 陆地棉AOS基因家族成员在不同组织中的表达量
Fig. 5 Expression levels of the AOS gene family in different tissues of G. hirsutum
从不同组织来看,在体细胞胚再生中的表达模式大致可分为3类:GhAOS1和GhAOS6在棉花胚发育过程中表达量相对较高;GhAOS2和GhAOS3在棉花愈伤和胚性愈伤中表达量相对较高;GhAOS4和GhAOS5在茎和叶中表达水平相对较高。其中GhAOS1在体细胞胚发育过程中表达量持续升高,且表达量明显高于其他基因;GhAOS2主要在愈伤和胚性愈伤中表达,从球形胚到子叶胚的发育阶段中表达水平比较低;而GhAOS4和GhAOS5基因的表达水平随着体细胞胚的发育表现出逐渐降低的趋势,与其他组织相比较,在体细胞胚中的表达水平显著降低;GhAOS3与GhAOS6基因转录组的结果和qRT-PCR结果有一定的差异。
AOS基因作为JA合成限速酶基因,在调控JA合成和信号转导方面发挥重要的作用。合子胚发育和体细胞胚发育过程高度相似,AOS通过调控JA水平影响种子的萌发和生长,所以笔者推测AOS在调控体细胞胚发育方面也发挥着重要的作用。本研究利用棉花体细胞胚发育转录组数据,找到6个AOS基因家族成员,发现这6个基因相似性较高,GhAOS2与GhAOS6可能是由同一个基因复制后分布到了不同染色体上;GhAOS3与亚洲棉叶绿体中的GaAOS在棉花进化过程中可能发挥相同的作用[15-17];GhAOS5与已经报道的GhAOS可能是同一个基因。6个基因中只有GhAOS1和GhAOS6有内含子,而且它们被定位在不同的染色体上,说明在基因的转录过程中发生了不同的加工过程,基因功能可能与它们在染色体的位置有一定的关系。基因亚细胞定位分析发现这些基因定位在不同的部位,说明它们可能行使不同的功能。GhAOS1、GhAOS6都在过氧化物酶体中起作用,而植物中过氧化物酶体功能主要有:(1)参与光呼吸作用;(2)在萌发的种子中,进行脂肪的β-氧化[26-27]。体细胞胚和合子胚发育高度相似,推测这些基因可能参与体细胞胚的光呼吸、萌发和成熟。这与它们的组织表达模式分析相一致,GhAOS1和GhAOS6在棉花体细胞胚中具有较高的表达水平,暗示这2个基因在棉花体细胞胚发育过程中发挥重要作用。GhAOS3被定位在叶绿体基质中,进化树分析也证明了它与陆地棉和亚洲棉的叶绿体中的AOS进化关系最近,推测其可能与体细胞胚发育过程中的光合作用有关。GhAOS2预测定位在线粒体基质内,暗示它可能参与了线粒体内的能量代谢过程。根据GhAOS2和GhAOS3的亚细胞定位,推测这2个基因可能与提供能量有关,而且它们的表达模式相似,都在胚性愈伤中的表达水平最高,说明这2个基因可能通过协同作用调控胚性愈伤分化产生体细胞胚。对AOS基因的启动子元件预测分析发现所有的AOS基因的启动子都含有SKn-1基序,表明它们都在胚发育中发挥一定的作用,由于它们表达水平的不同,其在调控胚发育方面发挥的作用大小也不同。GhAOS2和GhAOS6都存在WUN基序,表明这2个基因受伤害诱导,这与植物受到机械损伤后,JA表达量增加的报道相似[28-29]。不同基因的启动子含有的顺式作用元件不尽相同,可能受到不同激素和不同胁迫的调控,在棉花体细胞胚发育过程中可能以协同或拮抗的方式发挥作用。
根据转录组数据和qRT-PCR结果,大部分AOS基因的表达模式是相似的,但GhAOS3与GhAOS6基因转录组的结果和qRT-PCR结果有一定的差异,其原因推测可能是由于转录组测序是打断mRNA测小片段的,后面需要拼接,但实际上由于偏好性的存在,某些片段会被多次测到,而某些片段被测到的概率低,尤其是对于相似性很高的基因,转录组数据可能会有一定的偏差。总的来讲,GhAOS3与GhAOS2可能以协同互作的方式调控胚性愈伤分化产生体细胞胚,GhAOS1和GhAOS6在棉花体细胞胚中的表达水平最高,而且随着体细胞胚的发育表达水平在逐渐增加,说明棉花体细胞胚发育过程中需要的JA含量在不断增加,外源茉莉酸处理促进棉花体细胞胚发育[30]。
合适的逆境条件有利于体细胞胚的发育,而AOS基因的启动子中有许多响应逆境胁迫的元件,因此作者推测胁迫条件下可能通过诱导AOS基因的表达促进体细胞胚胎发生;转录组数据和qRT-PCR结果也证明不同的AOS基因有时空表达特异性,在调控棉花体细胞胚发育方面发挥不同的作用。这与染色体定位,亚细胞定位和启动子分析结果相一致,即GhAOS基因家族在陆地棉体细胞发育中发挥重要的作用,但不同的AOS基因的作用不尽相同。通过对AOS基因的染色体位置、亚细胞定位、启动子和表达模式分析,有效地预测出AOS基因主要在哪个发育过程发挥作用,筛选出主要在棉花体细胞胚发育过程中发挥主要作用的AOS基因,为下一步系统研究AOS基因调控棉花体细胞胚胎发生的机制奠定了理论基础。
The authors have declared that no competing interests exist.
| [1] |
Five genera of Magnoliaceae occur in China; these and the 81 indigenous and 7 cultivated species they contain are described here. In Magnolia, 28 wild and 4 cultivated species are recognized. Magnolia nitida var. robusta is newly described, and Magnolia lotungensis is reduced to Magnolia nitida var. lotungensis. In Manglietia, 17 indigenous and 1 cultivated species are recognized. Section Manglietiastrum is transferred from Magnolia to Manglietia, and Magnolia sinica is renamed Manglietia sinica. Manglietia forrestii is reduced to Manglietia fordiana var. forrestii, Manglietia kwangtungensis is reduced to Manglietia fordiana var. kwangtungensis, and Manglietia calcarea is reduced to Manglietia fordiana var. calcarea. In Kmeria, one species is recorded for China. In Michelia, 34 indigenous and 1 cultivated species are recognized. Section Tsoongiodendron, based on Tsoongiodendron W. Y. Chun, is recognized with one species, Michelia odora; section Paramichelia, based on Paramichelia H. H. Hu, is recognized with one species in China. Michelia elliptilimba is newly described. Michelia crassipes is reduced to Michelia figo var. crassipes. In Liriodendron one indigenous and one cultivated species are recognized. An identification list of all the collections studied, as well as a bibliography of Chinese Magnoliaceae, is given at the end of the paper.
|
| [2] |
[本文引用:1]
|
| [3] |
<a name="Abs1"></a>Plant responses to wounding and herbivore attack are orchestrated by complex signaling pathways that link the production of chemical and physical signals at the wound site to activation of gene expression and other cellular processes. The systemic nature of many wound-induced responses provides an attractive opportunity to study intercellular signaling pathways that operate over long distances within the plant. Genetic dissection of the wound-response pathway in tomato indicates that (1) systemin and its precursor protein, prosystemin, are upstream components of an intercellular signaling cascade that requires the biosynthesis and action of jasmonic acid (JA); and (2) physiological processes regulated by this pathway confer host resistance to a broad spectrum of plant invaders. Grafting experiments conducted with mutants defective in systemic wound signaling indicate that systemin functions at or near the wound site to trigger the production of JA, which in turn acts non-cell autonomously to promote systemic defense responses. The location of JA biosynthetic enzymes within the companion cell-sieve element complex of vascular bundles, together with the accumulation of JA in vascular tissues, support a role for jasmonates as phloem-mobile signals. The recent discovery of enzymes involved in the metabolism of JA to volatile methyl-JA and bioactive JA-amino acid conjugates has potential implications for the mechanism by which JA promotes wound signaling. Species-specific differences in the mechanism of wound signaling appear to reflect the way in which the wound-induced jasmonate pathway is regulated by other signals including systemin, cell wall-derived oligosaccharides, ethylene, and insect-derived elicitors. Adding to the complexity of the wound-induced jasmonate cascade are wound-signaling pathways that operate independently of JA.
|
| [4] |
Plants respond to mechanical wounding or herbivore attack with a complex scenario of sequential, antagonistic or synergistic action of different signals leading to defense gene expression. Tomato plants were used as a model system since the peptide systemin and the lipid-derived jasmonic acid (JA) were recognized as essential signals in wound-induced gene expression. In this review recent data are discussed with emphasis on wound-signaling in tomato. The following aspects are covered: (i) systemin signaling, (ii) JA biosynthesis and action, (iii) orchestration of various signals such as JA, H2O2, NO, and salicylate, (iv) local and systemic response, and (v) amplification in wound signaling. The common occurrence of JA biosynthesis and systemin generation in the vascular bundles suggest JA as the systemic signal. Grafting experiments with JA-deficient, JA-insensitive and systemin-insensitive mutants strongly support this assumption.
|
| [5] |
早衰是棉花生长发育的一种异常现象,是大量衰老相关基因差异表达 的结果.早衰棉花光合作用、碳水化合物和其他生物大分子合成相关基因大多下调表达,而蛋白、核苷酸、脂类降解和氨基酸、糖类、嘌呤、嘧啶和离子转运体等养 分循环利用相关基因大多上调表达;脱落酸(ABA)、乙烯、生长素、茉莉酸(JA)和赤霉素(GA)相关基因大多上调表达,而细胞分裂素合成基因IPT下 调表达;NAC和WRKY等转录因子基因也大多上调表达.结合作者在该领域的研究,总结评述了光合作用及大分子降解、养分循环利用、激素和转录因子相关基 因在早衰棉花中的表达模式及作用机理.
URL
[本文引用:]
|
| [6] |
植物中庞大的AP2基因家族成员因其广泛参与植物响应外界环境胁 迫、生长发育相关的转录调控而备受重视.AP2基因曾被认为植物所特有,但最近在蓝藻、线虫和病毒中发现了具有AP2结构域和位点特异核酸内切酶的蛋白. 所以有人认为当今植物中的AP2基因起源于细菌或者病毒的基因的横向转移,AP2结构域可能来自后来进化为叶绿体的原始蓝细菌的内共生.ERF是AP2大 家族中的一个亚族,它编码的蛋白能特异结合含有GCC盒的病程相关基因的表达,参与植物抗病反应.ERF基因的表达受到疾病相关刺激以及环境胁迫的诱导, 并且在乙烯、茉莉酸和水杨酸信号传导途径中发挥一定的作用.同时,某些ERF基因在转基因植物中的超表达表现了一定的广谱抗性,因而在分子育种中具有一定 的应用前景.棉花上AP2基因家族的基因克隆与分析最近才得以进行,介绍了我们在棉花上相关的研究工作并讨论了它们在植物抗病反应中的作用.
|
| [7] |
Jasmonates are a new group of plant hormones; their roles on plant development are still little known. The aim of this work is to determine the action of jasmonates on cabbage, Brassica oleracea L. var Capitata , development both in in vitro cultured explants and in whole plants. Jasmonic acid (JA) enhanced nodal explant development when applied at 2–50 nM and inhibited it when supplied at 1250 and 6000 nM JA. Overall plant development was enhanced most under the 10 nM JA treatment; which significantly increased the explant shoot, leaf, and root dry weight. The root system of the explants cultured under the lower JA concentrations appeared more vigorous. Jasmonic acid also promoted the development of isolated in vitro cultured roots when applied at 2 and 10 nM . Root length and weight significantly increased, while concentrations 250 nM JA and over were detrimental. Isolated roots were progressively thicker as the JA concentration increased. Methyl jasmonate promoted both the below- and above-ground cabbage plant development when applied in a confined atmosphere at a concentration of only 1.225 nl.l 611 MJ: plants were higher and heavier, and showed an improved root system development. On the other hand, the 2.43 nl.l 611 MJ treatment decreased plant growth. The present work reveals a role for jasmonates as enhancers of in vitro and in vivo cabbage plant development. To our knowledge, no corresponding studies on the effects of jasmonates on whole plants have been previously published.
|
| [8] |
Abstract Jasmonic acid (JA) is implicated in a wide variety of developmental and physiological processes in plants. Here, we studied the effects of JA and the combination of JA and ethylenediamine-dio-hydroxyphenyl-acetic acid (EDDHA) on flowering in Lemna minor in axenical cultures. JA (0.475-47.5 nmol l(-1)) enhanced floral induction in L. minor under long-day (LD) conditions. Under the same conditions, at a concentration of 237.5 nmol l(-1), JA inhibited floral induction, and at a concentration of 475 nmol l(-1) it prevented floral induction. Under LD conditions with LD preculture, a combination of EDDHA (20,500 nmol l(-1)) and JA (47.5 nmol l(-1)) had a synergistic effect on the promotion of floral induction. Floral induction was enhanced to the greatest extent in experiments with LD precultures. Microscopic examination of microphotographs of histological sections showed that JA and, to an even greater extent, JA+EDDHA at optimal concentrations promote apical floral induction (evocation). Furthermore, JA, and to an even greater extent JA in combination with EDDHA in an optimal concentration, also promote flower differentiation, especially the development of stamens, as is evident from the microphotographs. The experimental results show that JA promotes floral induction in other species of Lemnaceae from various groups according to their photoperiodic response. The results support our hypothesis that, in addition to previously ascribed functions, JA may regulate floral induction, evocation and floral differentiation. Our hypothesis is supported also by the results obtained by quantitative determination of endogenous JA levels in L. minor at three growth stages. The levels of endogenous JA decreased from 389 ng JA g(-1) (fresh weight) of L. minor during the vegetative stage to 217 ng JA g(-1) during the evocation stage, and to 37.5 ng JA g(-1) during the flowering stage, which proves that JA is used for flowering.
|
| [9] |
URL
[本文引用:1]
|
| [10] |
[本文引用:1]
|
| [11] |
[本文引用:1]
|
| [12] |
URL
[本文引用:1]
|
| [13] |
[本文引用:1]
|
| [14] |
The octadecanoid pathway metabolite jasmonic acid (JA) plays a vital role in rice (Oryza sativa L. cv. Nipponbare) defense/stress response(s). However, genes involved in its biosynthesis remain unidentified. Here, we cloned a novel rice cDNA highly homologous to the allene oxide synthase (EC 4.2.1.92) AOS gene, the first committed step in JA biosynthesis, showing significant similarity at the amino acid level with a related monocotyledoneous barley AOS. OsAOS is a novel member of the cytochrome P450 CYP74A subfamily and exists as a single copy gene in the rice genome. An examination of its steady state mRNA level in two-week-old seedling leaves revealed that OsAOS does not express constitutively in healthy leaves, and shows a weak responsiveness to cut. Signaling components of defense/stress pathways, in particular JA itself, strongly up-regulated the OsAOS transcript, whereas salicylate, ethylene, abscisic acid, and hydrogen peroxide were not so effective. Copper, a heavy metal also significantly enhanced the OsAOS expression. Protein phosphatase inhibitors proved to be the most potent in up-regulating the OsAOS mRNA level, suggesting the involvement of phosphorylation/dephosphorylation events in its regulation. Moreover, the inducible nature of OsAOS was influenced by light signal(s). Blast pathogen (Magnaporthe grisea) specifically elicited the accumulation of OsAOS mRNA in leaves in an incompatible versus compatible interaction, a first demonstration of pathogen responsiveness for any AOS gene to date. These results strongly suggest the importance of OsAOS in rice defense/stress response pathway(s).
|
| [15] |
The gene encoding rice allene oxide synthase, OsAOS, was intronless and had nucleotide sequences with the high GC content of 67%. Deduced amino acid sequences had very high similarity with other AOS proteins, in particular 74% similarity to barley, characterized by the conserved motifs of P450 cytochrome of the CYP74A family. Purified recombinant rice AOS protein expressed in Escherichia coli converted 13-hydroperoxylinolenic acid to allene oxide. Several restriction enzyme digestions and Southern analysis showed that OsAOS was likely to have two copies in its genome. The basal level of OsAOS expression was detected in various tissues and the transcription level was increased by jasmonate treatment.
|
| [16] |
Abstract The Arabidopsis mutant defective in anther dehiscence1 (dad1) shows defects in anther dehiscence, pollen maturation, and flower opening. The defects were rescued by the exogenous application of jasmonic acid (JA) or linolenic acid, which is consistent with the reduced accumulation of JA in the dad1 flower buds. We identified the DAD1 gene by T-DNA tagging, which is characteristic to a putative N-terminal transit peptide and a conserved motif found in lipase active sites. DAD1 protein expressed in Escherichia coli hydrolyzed phospholipids in an sn-1-specific manner, and DAD1-green fluorescent protein fusion protein expressed in leaf epidermal cells localized predominantly in chloroplasts. These results indicate that the DAD1 protein is a chloroplastic phospholipase A1 that catalyzes the initial step of JA biosynthesis. DAD1 promoter::beta-glucuronidase analysis revealed that the expression of DAD1 is restricted in the stamen filaments. A model is presented in which JA synthesized in the filaments regulates the water transport in stamens and petals.
|
| [17] |
[本文引用:2]
|
| [18] |
In this review, we address the role of stress as one of the principal causes for a cell or tissue to change its preexisting somatic program, reprogramming itself to express the embryogenic pathway. The focus of this paper is the effect of different stress conditions on the induction phase of plant somatic embryogenesis, as well as the development of embryogenic competence as a result of the applied stresses. We also present a variety of data that link plant somatic embryogenesis, DNA methylation and oxidative stress response.
|
| [19] |
Gossypium hirsutum has proven difficult to sequence owing to its complex allotetraploid (AtDt) genome. Here we produce a draft genome using 181-fold paired-end sequences assisted by fivefold BAC-to-BAC sequences and a high-resolution genetic map. In our assembly 88.5% of the 2,173-Mb scaffolds, which cover 89.6%鈭96.7% of the AtDt genome, are anchored and oriented to 26 pseudochromosomes. Comparison of this G. hirsutum AtDt genome with the already sequenced diploid Gossypium arboreum (AA) and Gossypium raimondii (DD) genomes revealed conserved gene order. Repeated sequences account for 67.2% of the AtDt genome, and transposable elements (TEs) originating from Dt seem more active than from At. Reduction in the AtDt genome size occurred after allopolyploidization. The A or At genome may have undergone positive selection for fiber traits. Concerted evolution of different regulatory mechanisms for Cellulose synthase (CesA) and 1-Aminocyclopropane-1-carboxylic acid oxidase1 and 3 (ACO1,3) may be important for enhanced fiber production in G. hirsutum.
|
| [20] |
Upland cotton is a model for polyploid crop domestication and transgenic improvement. Here we sequenced the allotetraploid Gossypium hirsutum L. acc. TM-1 genome by integrating whole-genome shotgun reads, bacterial artificial chromosome (BAC)-end sequences and genotype-by-sequencing genetic maps. We assembled and annotated 32,032 A-subgenome genes and 34,402 D-subgenome genes. Structural rearrangements, gene loss, disrupted genes and sequence divergence were more common in the A subgenome than in the D subgenome, suggesting asymmetric evolution. However, no genome-wide expression dominance was found between the subgenomes. Genomic signatures of selection and domestication are associated with positively selected genes (PSGs) for fiber improvement in the A subgenome and for stress tolerance in the D subgenome. This draft genome sequence provides a resource for engineering superior cotton lines.
|
| [21] |
过氧化物酶体参与了包括氧化氢反应、长链脂肪酸的β-氧化等几乎所有的必需代谢途径.植物过氧化物酶体在植物体抗病和抗衰老过程中发挥作用.介绍了植物过 氧化物酶体与亚硫酸盐氧化酶以及植物过氧化物酶体抗衰老、生物发生和动力学等方面的研究进展.
|
| [22] |
[本文引用:]
|
| [23] |
[本文引用:]
|
| [24] |
URL
[本文引用:]
|
| [25] |
[本文引用:]
|


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}